A dolgozat I. részében a liszteskék (Aleyrodidae) családjáról szóló fontos ismereteket gyűjtöttük össze. A II. részben az aukubaliszteske megismerését megalapozó – 2020 és 2023 őszén az „A” vizsgálati helyen folytatott helyszíni és „házi-laboratóriumi” megfigyeléseinkről számolunk be. Ugyanebben a részben soroltuk fel a 2024. augusztus-szeptemberi vizuális szemléink során észlelt - hazánkban és külföldön még nem jelzett - gazdanövényeket is. A dolgozat III. (jelen) részében a 2024. április közepe-október közepe időszakban végzett vizsgálatainkról tájékoztatjuk az Olvasókat.
(Fontos megjegyzés: a dolgozatban a ’bölcső’ és a ’pupárium’ kifejezéseket a ’pszeudopupa’ értelemben, annak stilisztikai helyettesítésére használjuk.)
A vizsgálatok – egymástól légvonalban kb. 200 m távolságra lévő – két helyszínen történtek. Ezekre a továbbiakban ’A’ vizsgálati hely és ’B’ vizsgálati hely neveken hivatkozunk.
Az aukubaliszteskék rossz repülők, saját erejükből nem képesek gazdanövényüktől nagyobb távolságra elrepülni. A gazdanövényen lerakott petékből kikelő L1 stádiumú lárva mozgásképes (többségük nem hagyja el azt a levelet, amelyen a petéből kikelt), a többi preimaginális fejlődési alak helyhez kötött. Az áttelelő stádium a pszeudopupa, amely lombhulláskor a levéllel együtt leesik.
A helyszínen is könnyen megtalálható pszeudopupák előfordulása és a levelenkénti denzitása szerint egyik legkedveltebb és elterjedt hazai gazdanövénye a nyugati ostorfa (Celtis occidentalis), ezért a részletes vizsgálatokat az erről a gazdanövényről gyűjtött levélmintákon végeztük.
Kipróbálásra szánt módszerünk tervezését a fenti ismeretekre alapoztuk, ezért ’A’ vizsgálati helyként a 2023 szeptemberében Budapest XI. kerületében erős pszeudopupa fertőzöttséget (esetenként 20 db bölcső/levél sűrűséget is számoltunk) mutató ostorfákat választottunk ki. Arra gondoltunk, hogy a nagyobb telelő népességből a következő év tavaszán több imágó kel ki, és így az első, ill. további nemzedék(ek) különböző fejlődési stádiumú egyedeit is könnyebben fogjuk megtalálni.
Mivel a fajról szóló, általunk olvasott dolgozatok és a közösségi média elsősorban a faj ’bölcsőit’ mutatják be, ezért törekedtünk arra, hogy a faj életciklusára és a faj fejlődési alakjaira vonatkozóan minél több megfigyelést végezzünk.
A begyűjtött levélmintákon talált fejlődési alakok nagyságát is megmértük. Mivel az imágók és a pszeudopupák kivételével a többi fejlődési alak szabad szemmel nem látható (méret és szín), ezért fontosnak tartottuk, hogy az egyes fejlődési alakokról készített képeinket is közreadjuk.
Mivel az egyes fejlődési alakok méretét más forrásokból nem ismerjük, ezért az általunk megadott méretek ellenőrzése/megerősítése kívánatos.
Az első – a levelek fonákán egyesével elhelyezkedő – imágókat 2024.05.06.án láttuk az ’A’ vizsgálati hely növényein. Az elhúzódó imágókelés miatt az észlelt egyedek száma lassan gyarapodott, de a 2024.05.16.-i helyszíni szemlénken már csoportosuló és párba álló és egyedeket is fényképeztünk. A begyűjtött és mikroszkóp alatt megvizsgált leveleken petéket és lárvákat még nem találtunk.
Mint a dolgozat I. részében említettük Kanmiya, K. (2005) 36 Japánban élő liszteske fajnál (köztük az Aleuroclava aucubae fajnál is) kimutatta, hogy a párkeresésben, a nemek egymásra találásában, az udvarlásban az egyedek közt vibrációs kommunikáció történik. Tanulmányának kézi rajzán ábrázolta a Trialeurodes veporariorum udvarlási viselkedésének ismétlődő (sztereotip) fázisainak sorrendjét.
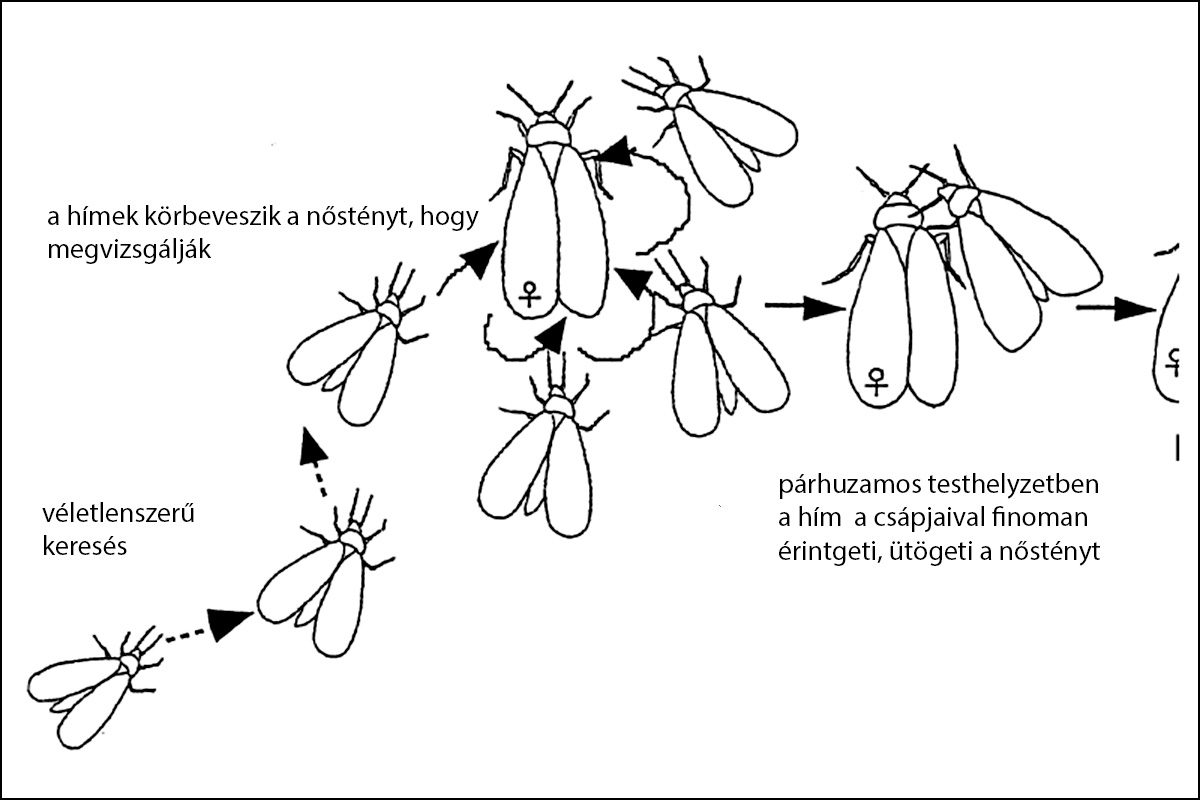
A fentiekben bemutatott udvarlási fázisokat az aukubaliszteske imágóinak helyszíni fényképezése során a tervezett ’A’ vizsgálati helyen több időpontban, (2024 május közepén, 2024.07.28., 2024.08.25., 2024.08.31.- én tömeges imágórajzást fényképeztünk.
Eredeti tervünkben rendszeres megfigyelésre további vizsgálati helyet nem kívántunk bevonni, de a közterületi zöldfelületek vizuális szemléi során egy vegyes fajú cserjefoltban magról nőtt fiatal ostorfa növényeken 2024.07.28.-án tömegesen rajzó imágókat láttunk. Emiatt a fentebb körülírt növényeket ’B’ vizsgálati hely megjelöléssel a továbbiakban rendszeresen szemléztük. (2024.07.28., 2024.08.06., 2024.10.09.)
A tipikus udvarlási fázisokat mindkét helyen megfigyeltük.

Az aukubaliszteskékre jellemző, hogy a hímek valamivel kisebbek a nőstényeknél. A méretbeli különbség az összes képrészleten jól látható.
Az 1/1. sz. képen az ostorfa levélfonákán gyülekező aukubaliszteske imágói láthatók.
Az 1/2-1/5. sz. képek a keresés és az udvarlás egyes fázisait mutatják:
Az 1/2. sz. képen véletlenszerű keresés, az 1/3. sz. kép: a csalogatott hímek körbeveszik a nőstényt, az 1/4. sz. képen egymással közel párhuzamos helyzetben a hím a csápjaival érintgeti/finoman ütögeti a nőstényt. Az 1/5. sz. képen párosodás közben a hím (jobbra) szárnyait ráhelyezi a nőstény szárnyaira Forrás: ITT.
A begyűjtött leveleken minden fejlődési alakot megtaláltunk.
Az imágók tömeges gyülekezését és párba állását figyeltük meg az ’A’ vizsgálati helyen, 2024.08.25.-én reggel 7 órakor 24 °C hőmérsékleten, teljes szélcsendben nyugati ostorfa fiatal hajtásvégein.

Az ’A’ vizsgálati hely fényképét lásd a 14/1 képen.
A korábbi helyszíni megfigyelések alkalmával az imágók csak zavarásra, a növény megmozdításakor keltek szárnyra, majd 1-2 mp-en belül leszálltak egy levél fonákára. Ebben az esetben viszont láttuk az imágók spontán felrepülését is, de ekkor sem távolodtak el a felszállási helytől 30-40 cm-nél távolabbra.
Közbevetőleg megjegyezzük, hogy 2024. augusztus-szeptember hónapokban a gazdanövénykör részletesebb megismerése céljából végzett helyszíni szemléink során egyetlen más gazdanövényen sem láttunk ilyen rendkívüli imágórajzást, ill. gyülekezést.
A képen láthatóhoz hasonló néhány fiatal hajtás 30-40 cm-es végszakaszát levágtuk, majd a rajtuk elhelyezkedő leveleket mikroszkóp alatt megvizsgáltuk. Összesen 60 db levelet gyűjtöttünk (a levélváll és a levélcsúcs távolsága alapján méret szerint csoportosítva: 30 db 8-12 cm hosszúságú, 17 db 6-8 cm-es és 13 db 4-6 cm-es levelet), amelyeken összesen 2 db petét és 1 db L1 mozgó lárvát találtunk. de egyetlen idősebb lárvát vagy pszeudopupát sem fedeztünk fel. A levélméret és a rajtuk talált fejlődési alakok között nem találtunk összefüggést.
Ugyanitt 1 héttel későbbi (2024.08.31.) az imágók rajzása még mindig tartott. A hajtásvégek hajtáscsúcstól számított 4.-6. levélemeletének begyűjtött levélmintáin (30 db) minden fejlődési alakból találtunk néhány egyedet.
A következő képeinken az egyes fejlődési alakokról készült felvételeinket mutatjuk be.

Az imágók olyan gyengén kitinizáltak, hogy a makrofotók készítése során a (dorzális-ventrális-laterális) testhelyzet beállításakor a puha szőrű ecset egyetlen sertéjétől is könnyen megsérültek. Ennek elkerülésére a levél ellentétes oldali ’kopogtatásának’ módszerét használtuk.
Méréseink szerint az imágók testhossza a szárnyakkal együtt 0,9-1,1 mm, szélessége 0,4-0,5 mm. A hímek valamivel (10-15%) kisebbek a nőstényeknél. A szárnyak egyöntetűen fehér viaszporral fedettek, rajtuk mintázat, foltok nem láthatók. Nyugalmi állapotban a szárnyakat a potroh felett háztetőszerűen tartják.
A fej és lábak sárgásfehérek, az összetett szemek sötétvörösek.
A hímeket és nőstényeket a potroh szélesség / hosszúság aránya alapján meg lehet különböztetni. A hímek potroha karcsúbb, a nőstényeké – a potrohban fejlődő peték miatt – zömökebb. A hímek potroha a jellegzetes oldalsó párosodási helyzet miatt is karcsúbb, hosszabb (lásd a 3/2 és a 3/4 sz. képeket).
A tor és a potroh sárgás-fehéres színű. A hímek párzószervét a nőstényhez való rögzülést szolgáló páros függelék (paramer) veszi körül. A nőstényeknek rövid tojócsövük van. Továbbiakat lásd a pete ismertetésénél.
Az 3/1-2 sz. képeken hím imágókat mutatunk dorzális és ventrális nézetben. A ventrális nézeten a karcsú potroh végén a paramer jól kivehető.
A 3/3.-4. sz. képek nőstényeket mutatnak laterális és ventrális nézetben. A laterális nézeten jól elkülönül a rövid tojócső.
Megjegyzés: Az 3/1-4 sz. képek nem azonos nagyításban mutatják az egyedeket (nem arányosak).
Számos liszteske-faj csoportosan, sokszor kör/ovális alakban vagy rendezetlenül rakja le a petéket, amelyeket fehér viaszporral fed be illetve a peték körül is fehéres viaszporos zóna látható.
Patel C., Srivastava R.M., Samraj J.M. (2022) ismertetik két világszerte fontos faj (Bemisia tabaci és a Trialeurodes vaporariorum) fejlődési stádiumainak összehasonlító morfológiai vizsgálatát. A petéknek a levél szövetébe történő elhelyezéséről megállapítja, hogy a nőstények a petéket a tojócső (ovipozitor) segítségével juttatják a levélfonák szövetébe.
Az ovipozitor az alsó epidermiszt átszúrva egy nyílást készít a pete alsó (szubterminális) végén lévő petenyél (pedicel) számára. A petenyél rögzítését részben a tojócső által szétfeszített szövetek rugalmassága, valamint a növényi turgornyomás biztosítja.
Penny J. Gullan, Jon H. Martin (2009) szerint az Aleyrodidae családra jellemző, hogy a pedicel a lárva kikeléséig nedvességet szív fel a levél szövetéből.
Az aukubaliszteske nőstényei a petéket egyesével helyezik el elsősorban a levelek fonákának érközi részein. A nőstények átlagos peteprodukcióját nem ismerjük.

Vizsgálataink során lefényképeztük az aukubaliszteske petéit és az alsó végükön elhelyezkedő petenyélnek a növényi szövetből kiálló részét. Méréseink szerint a peték hosszúsága X szélessége: 0,18-0,2 mm X 0,04-0,06 mm nagyságúak.
A peték többnyire ferdén (nem merőlegesen) állnak a levélfonák síkjára, annak érközi részein elszórtan, egyesével helyezkednek el, rajtuk és környező levélfelületeken viasz-szemcséket nem láttunk. Alakjuk ovális, felső végük kissé csúcsosabb, legszélesebb részük a felső véghez közelebb található. A frissen lerakott peték fehéres áttetszőek, alsó és felső csúcsuk az érés során sárgásra színeződik.
A 4/1. sz. képen egy párban álló hímet (felül) és egy nőstényt, valamint a jobb felső sarokban egy frissen lerakott fehéres/áttetsző petét látunk.
A 4/2. sz. kép egy fekete pszeudopupát és a felső sarkokban 1-1 oldalán felrepedt üres petehéjat mutatunk.
Az említett képeket az imágók:pete és a pszeudopupa:pete méretarányainak bemutatására is szánjuk.
A 4/3/a sz. kép egy frissen lerakott, majdnem fehér pete, a 4/3/b képen néhány napos peték, a 4/3/c képen üres petehéjak láthatók. Az L1-es lárva kelésekor a petehéj az oldalán reped fel.
A 4/4. sz. képen részletesebben, két irányból szemléltetünk egy-egy petét.
Az a.) részleten a pete dorzális felületét látjuk, amelynek a pedicellel (petenyél) szemben lévő sárga csúcs alatt kétoldalt egy-egy pirosas szemfolt található, itt fejlődnek ki az L1-es mozgásképes lárva szemei. A szemfoltok elhelyezkedése egyben meghatározza a pete elülső (anterior) csúcsát és a háti (dorzális) felületet is.
A b.) részleten egy másik petét mutatunk, a jobbra eső domború része a dorzális- (háti), vele átellenesen pedig a – majdnem sík – ventrális (hasi) felülete látható oldalnézetben. A két nézet összevetése alapján a peték hosszirányra merőleges keresztmetszete egyik oldalán lapos, félkör alakú.
Mint a dolgozat I. részében említettük a liszteskék posztembrionális fejlődése az epimorfózis speciális formája, az allometabólia. A nyéllel a levélhez rögzített petéből kikelő lárva mozgásképes, lábai és csápjai viszonylag hosszúak.
Az 1. fokozatú lárvák keresik (ez több óra is lehet) a megtelepedésre alkalmas helyet, ahol szipókájukat a levélszövetébe mélyesztve végleg megtelepednek és helyhez kötött lárvává alakulnak. A többi (2-4.) stádiumban a fejlődési alakok lábai, csápjai igen rövidek, szárnykezdeményeik nem fejlődnek ki. Az utolsó (4.) stádiumú nyugalmi állapotú úgynevezett pszeudopupából (korábban lárvabölcső, tévesen pupárium) bújik ki az imágó.

Az 5/1-4. sz. képeken egy L1-es lárva mozgását követtük és néhány mp-es időközönként lefényképeztük: 1: 11:38:30; 2. 11:38:42; 3: 11:38:50; 4: 11:38:58. A sorszámozott képek az óramutató járásával megegyező irányú sorrendben mutatja be a lárva pillanatnyi helyzetét. A mozgás jobb követhetősége érdekében a digitális felvételen utólag piros színnel megjelöltük a levél egy rögzített viszonyítási pontját.
A lárva testtájainak könnyebb azonosításához javasoljuk figyelembe venni, hogy a lárva háti felületének hosszanti középvonalában – a test felétől a végbélig – egy rövid zöldes vonal látható. Értelemszerűen ezzel szemben helyeződik az elülső (anterior) vagy feji testvég.
Ez a hosszanti szklerotizált felület az L2-L3 stádiumokban is megfigyelhető, de különösen jól látható a pszeudopupán. Gabrilov-Zimin, I.A., & Borisov, B. A. (2020) a pszeudopupa potrohának hátlemezén ezt a jellegzetes alakú részt ’trilobita-szerű’-nek nevezte. (A trilobiták kihalt háromkaréjú ősrákok.)
A fenti belső képek találkozásánál (középen) elhelyeztünk egy nagyobb felbontású képet az L1-es lárváról. Az L1-es lárvák nagyságát (hosszúság X szélesség) 0,24 X 0,14 mm-nek mértük. A lárvák lapítottak, a levéllemez síkjából alig emelkednek ki, alakjuk ovális, a tor és a potroh hátlemezének találkozásánál a legszélesebbek, a feji testvégen (a képen jobbra) szélesebben, az anális testvégen keskenyebben lekerekített. Az anális testvégen két farokserte helyezkedik el. A test pereménél kissé beljebb (szubmarginálisan) fehéres viasz-sáv látható.

Az egyes stádiumok a testméretek alapján is elkülöníthetők.
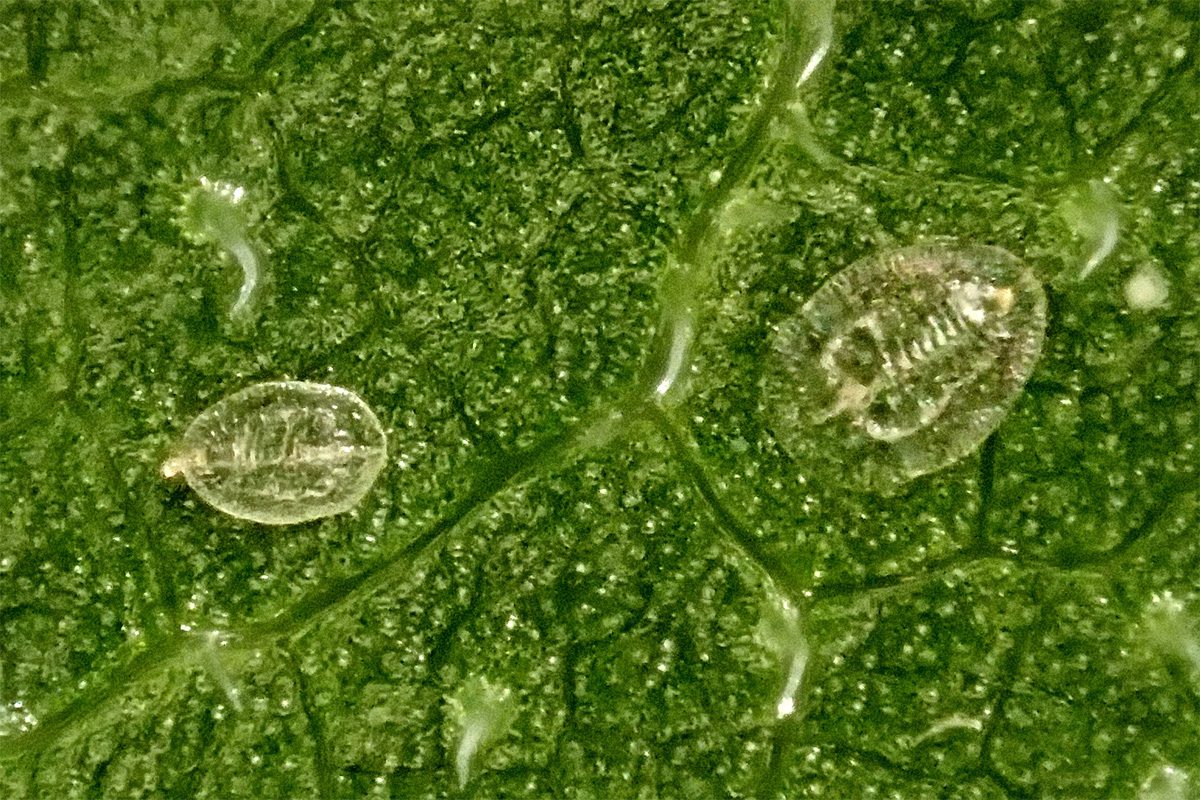
A képen balra egy L2-, jobbra pedig egy L3 stádiumú lárva látható. Nagyságukat (hosszúság X szélesség) L2: 0,35 mm X 0,23 mm (balra), L3: 0,5 x 0,35 mm-nek mértük. Alakjuk hasonló az L1 lárváéhoz, színük fehéres/áttetsző. Az L3 lárva viasz-szegélye arányaiban szélesebbnek látszik és a pszeudopupákra jellemző dorzális mintázat sokkal kifejezettebb.
A szúró-szívó szájszerv által helyhez kötött lárvák azonban nem mozdulatlanok. Szerencsés véletlennek köszönhető, hogy sikerült néhány képet készíteni egy L3-as lárva szipóka körüli forgó mozgásáról, melynek során a potrohát két alkalommal is felemeli.

A mozgássor 23 mp alatt lezajlott, ez idő alatt a lárva hossztengelye 28°-os szögben fordult el. A képeket az óramutató járásával egyező sorrendben egy gyűjtőképen mutatjuk be. A felvételek készítése során a fényképezőgép és a levél egymáshoz viszonyított helyzete változatlan volt.
A lárva elülső – szélesen lekerekített – testvége balra, a nyugati ostorfa levélfonáki szőrözött levélere közvetlen közelében látható. A lárvának az átellenes, keskenyen lekerekített testvégén két faroksertéje van. Annak érdekében, hogy az elfordulás jól érzékelhető legyen, ezért a képeken utólag piros vonallal megjelöltük a lárva hossztengelyét, és szoftveres támogatással meghatároztuk a hossztengelynek és a kép vízszintes tengelyével bezárt aktuális szögét is.
Az 8/1. sz. képen az induló állapot (12:01:58 mp) a hossztengely vízszintessel bezárt szöge 8°.
A 8/2. sz. képen (12:02:03 mp) a hossztengely vízszintessel bezárt szöge 25°. Egyben megjegyezzük, hogy a közelképekre jellemző nagyon kicsi mélységélesség miatt potroh felemelése elmosódottan látszik.
A 8/3. sz. képen (12:02:06 mp) a hossztengely vízszintessel bezárt szöge 16°.
A 8/4. sz. képen (12:02:25 mp) a hossztengely vízszintessel bezárt szöge -15°. A 8/3.-4. sz képek között (12:02:21-kor) készítettünk még egy felvételt, a mely a 2. sz. képhez hasonló potrohemelést mutatott, a -15°-os elfordulásnál. (ez a kép itt nem látható.)
A pszeudopupa fekete színe fokozatosan alakul ki.

A frissen kialakult pszeudopupák sárgászöldek, kissé áttetszőek, majd fokozatosan sötétednek és a folyamat végén teljesen megfeketednek. Rajtuk jellegzetes fehér viaszos mintázat jelenhet meg, de ez hiányozhat is.


A pszeudopupák rendkívül laposak, alig emelkednek ki a levélfonák síkjából. Hasi felületükkel szorosan illeszkednek a levél fonákához, dorzális nézetben alakjuk ovális.
A 11/1-2. kép Pyracantha coccinea leveléről készült úgy, hogy a levelet a fényképezés előtt a felső (adaxiális) levélfelület irányába ’hajlítottuk össze’, hogy a keletkező ’él’ megközelítően a pszeudopupa alatt helyezkedjen el.
A 11/3. képet fehér eperfa (Morus alba) leveléről készítettük. A levelet a pszeudopupa hossztengelyével párhuzamosan, a levélsíkra merőlegesen elvágtuk, majd a metszett felület felől fényképeztünk. A pszeudopupa faroksertéi mindegyik belső képen balra helyeződnek.
A pszeudopupa hosszirányú oldalnézetén látható, hogy a hasi felület sík, a hátlemez domború, leginkább kiemelkedő része a potroh középvonalában elhelyezkedő dudoroknál található, amely körben a perem felé folyamatosan vékonyodik. A perem függőleges széle ’recézett’-nek látszik.
Az Aleyrodidae család fajainak nőstény imágói általában nagyobbak a kifejlett hím egyedeknél. Lásd ITT. Az ivari dimorfizmust a 3. számú gyűjtőképen szemléltettük, és a képmagyarázatban részletes leírást is mellékeltünk.
Az áttanulmányozott irodalmakban csak két fajnál (Bemisia tabaci és Trialeurodes vaporariorum) találtunk utalást arra, hogy a nőstény pszeudopupák nagyobbak mint a hím ’bölcsők’ Bethke,J. A.; Paine, T. D.; Nuessly, G. S. (1991).
Mivel az egyes fejlődési stádiumok méretét a faji tulajdonságokon kívül a tápnövény és a mikrokörnyezet egyaránt befolyásolhatja, ezért olyan gazdanövény levelet kerestünk, ahol a különböző méretű pszeudopupák/exuviumok egymás mellett helyezkedtek el. Ezáltal ki lehetett zárni a tápnövény és a mikrokörnyezet különbözőségének hatásait.

A pszeudopupák közötti méretkülönbség a képen jól kivehető, azonban a nemi hovatartozást nem lehetett megállapítani, és ilyen jellegű irodalmi adatokról sem tudunk.
A lefényképezett pszeudopupák utólagos vizuális elemzése során feltűnt, hogy vannak közöttük kerekded és hosszúkásabb (karcsúbb) formájú egyedek is.
A keresztirányú vedlési varratnál mért legnagyobb szélességet összehasonlítottuk a hosszmérettel és ezt az arányszámot (mértékegység nélküli mérőszámot az ovális alak után) ovalitásnak neveztük el.
A 32 db fénykép alapján számítással meghatározott ovalitás értékek az alábbiak szerint összesíthetők:
| Ovalitás értéke | DB | % |
| <1.5 | 7 | 21,8 |
| ≥ 1.5 és ≤ 1.6 | 20 | 62,5 |
| >1.6 | 5 | 15,7 |
Mint korábban említettük a ’B’ vizsgálati helyet a 2024.07.28.-án észlelt intenzív imágórajzás miatt vettük fel a rendszeres megfigyelések közé. A vegyes összetételű cserjefoltban magról nőtt fiatal ostorfa növényeken az imágórajzás még 2024. 08.06.-án is folytatódott.
A 2024. augusztus 25-én ide tervezett helyszíni szemlénket sajnálatos módon nem tudtuk megtartani, mert a zöldterület fenntartói csonkolták a cserjecsoport fölé emelkedő ostorfa-vesszőket és hajtásokat, így hosszú időre lehetetlenné tették a további megfigyeléseket.
A vizsgálatsorozat lezárását jelentő 2024.10.09.-i helyszíni szemlénk során – az előzmények ellenére – ismételten felkerestük az említett növényeket, majd ezt követően az ’A’ vizsgálati helyet is.

A 13/1 kép áttekintő jelleggel távolabbról mutatja a cserjecsoportot. A befoglaló – részletesebb – kép középső részére mutató fehér vonal az ostorfák csonkolás után fejlődött hajtásainak helyét azonosítja. A hajtáscsúcsokon fejlődő levelek még nem érték el végleges nagyságukat.
A 13/2 képen két különböző levél fonákán fényképezett párban álló imágók láthatók. A kifejlett egyedek korábban már említett ivari dimorfizmusa (a hímek kisebbek) szabad szemmel is jól észlelhető.
A helyszíni fényképezés után gyűjtött levélmintákat ’házi-laboratóriumunkban’ mikroszkóp alatt megvizsgáltuk. A levelek fonákán találtunk frissen rakott (majdnem fehér) és néhány napos (sárgás) petéket és üres (oldalukon felrepedt) petehéjakat és L1 és L2 lárvákat. A petéket a 4/3/a-c képeken – a 2024.07.28.-i vizsgálatok alapján – korábban már bemutattuk. Kiemeljük, hogy L3-as lárvákat, pszeudopupákat, és az imágók kelése után visszamaradó exuviumokat nem találtunk.

A levelek mikroszkópos vizsgálata során egy felrepedt üres petehéj mellett mozgó frissen kelt L1-es lárvát láttunk. Az üres petehéj a képen az 5. szám felett található. A lárva mozgását 9 db önálló digitális képen rögzítettük, majd egy speciális képkezelési eljárással egy képpé montíroztuk.
A montírozás révén a helyváltoztatás fényképezett fázisai egyszerre láthatók. A rögzítés kezdetét az 1., az utolsó fázist pedig a 9. számmal jelöltük. Az 1. és a 9. képek készítése között 80 mp telt el. A képsorozat készítését követően is figyeltük a lárva mozgását. A mozgás nem volt folyamatos, a lárva helyenként néhány mp-re megállt és több alkalommal – sikertelenül – próbált átmászni a levél fonákán kiemelkedő ereken.
Külön nem jelöltük a kép bal alsó sarkában látható L2-es stádiumú lárvát.

Az ’A’ vizsgálati hely növényein új hajtásvégi levelek már nem fejlődtek, a növekedés leállt, a hajtások és a levelek a természetes őszi állapotukat mutatták 14/1-4 képek. A helyszínen kézi nagyítóval fekete pszeudopupákat és exuviumokat, a mikroszkópi vizsgálatok során pedig még néhány L3-as stádiumú lárvát és az elszíneződés előtt álló pszeudopupát találtunk.

A 15/1-2 képeken L3 stádiumú lárvák, a 15/3 képen elszíneződés előtti pszeudopupa látható. Külön említjük, hogy ismételten találtunk (lásd 15/4 képet) a 4 évvel ezelőtt (2020.10.10.) is észlelt gombával fertőzött pszeudopupákat. A korábbi felvételeket lásd a cikk II. részének 4. képén. Elérhető ITT.
Az ‘A’ és ‘B’ vizsgálati helyszíneken szerzett megfigyelések összegzése alapján arra a következtetésre juthatunk, hogy az imágók az ostorfán történő peterakás során kifejezetten előnyben részesítik a fiatal leveleket. Ezt a preferenciát már korábban is észleltük, amikor a párosodó imágók a hajtásvégi leveleken gyülekeztek. Lásd a 2. sz. képet.
A 2024.07.28.-án, a 2024.08.25.-én, a 2024.08.31-én és a 2024.10.09-én is észlelt kiugró imágórajzások – évjárattól és növényfajtól is függően – több, egymásba folyó nemzedékre utalnak, de az évenkénti nemzedékszám tisztázásához további vizsgálatok szükségesek.
Az egy nemzedék kifejlődéséhez szükséges idő a tápnövénytől és a hőmérséklettől is függhet, de ilyen vizsgálatokat nem végeztünk és irodalmi adatokról sem tudunk.
Összegzés:
Az aukubaliszteske pszeudopupáinak első hazai megjelenését 2014-ben észlelték néhány fás gazdanövényen Hári, K. et al. (2021).
A Budapest XI. kerületében 2024 augusztus-szeptemberben végzett gazdanövény-felderítéseink során a pszeudopupákat 26 db, nemzetközi kitekintésben is új gazdanövényen azonosítottuk. Az azonosításhoz nem a ’bölcsők’-nek az irodalomban kizárólagosan ajánlott részletes anatómiai vizsgálatát alkalmaztuk, hanem azok jellegzetes morfológiai bélyegeit hasonlítottuk össze az egyes gazdanövények pszeudopupáiról készült makrofotókon. Megjegyezzük, hogy hasonló méretű, színű és mintázatú pszeudopupákkal rendelkező liszteskék hazai előfordulásáról nincs tudomásunk.
A hazai gazdanövények száma az első híradásban felsorolt 10-ről 43-ra nőtt (külön számolva egyes fajok fajtáit és a hibrideket is). A pszeudopupák növényenkénti előfordulása és a levelenkénti denzitása alapján a gazdanövények nem azonos ’értékűek’, mert az egymástól néhány méter távolságban lévő gazdanövényeken lényegesen eltérő gyakoriságot tapasztaltunk.
Véleményünk szerint az aukubaliszteske legkedveltebb és vizsgálati területünkön (Budapest, XI. ker.) nagyon gyakori gazdanövénye a nyugati ostorfa (Celtis occidentalis).
Az eddig közterületi zöldfelületeken és a Budai Arborétumban talált gazdanövényeken a faj egyedeinek szívogatása kárt nem okozott, ellenük a kémiai védekezés nem indokolt.
A liszteske rejtőzködő életmódja és kicsinysége miatt a természetben a legtöbb gazdanövényen nehezen észlelhető ezért a fejlődési stádiumok vizsgálatát célzó helyszíni és ’házi laboratóriumi’ megfigyeléseinket a nyugati ostorfára (és annak begyűjtött leveleire) összpontosítottuk.
A beltéri izolált ’nevelési’ kísérletekhez rövid idejű (max. 3 nap) ún. ’megfigyelő dobozokat’ használtunk. Munkánkat nagyszámú helyszíni és makrofotóval dokumentáltuk (összesen 3559 db fényképet készítettünk), melyek közül néhányat dolgozatunkban is közzéteszünk.
A posztembrionális fejlődés egyes stádiumainak mikroszkópi megfigyelését és mérését (2020, 2023 őszén, valamint 2024. május-október időszakban) ’A’ és ’B’ vel jelölt vizsgálati helyein nőtt ostorfa növényeken végeztük.
Lefényképeztük a hím és nőstény imágókat (a potroh morfológiai összehasonlítása), a peték dorzális és laterális nézetét. Két különböző módszert dolgoztunk ki az L1-es lárvák levélfonáki mozgásának bemutatására, és gyűjtőképen tettük láthatóvá a helyváltoztatásra képtelen L3-as lárvák eddig nem ismert oldalirányú korlátozott ’forgását-rotációját’.
Irodalmi adatok hiányában szükségesnek tartjuk az általunk megadott méretek ellenőrzését.
Kipróbálásra szánt módszerünkben a helyszíni vizuális szemlék során az első imágók megjelenését 2024. május 6-án láttuk. Az áttelelő nemzedék imágóinak párosodására utaló udvarlási formákat (pl. hímek és nőstények szoros párba állása) 2024. május 15-én észleltük.
Ugyanilyen tömeges gyülekezést és viselkedési formákat észleltünk cserje jellegű közterületi nyugati ostorfák levélfonákán 2024. július 27-én, valamint 2024. augusztus 25-én és 31-én, míg 2024.09.07-én csak néhány (levélfonákon megbújó) imágót találtunk.
A vizsgálatsorozatot lezáró 2024.10.09-i helyszíni szemlénk során a ’B’ vizsgálati helyen, az augusztusban csonkára vágott fiatal ostorfa növények új hajtásain számos hím+nőstény párost fényképeztünk. A mikroszkópi vizsgálatok egy új nemzedék indulását mutatták (peték, L1-L2 lárvák).
Bizonytalan, hogy ennek a generációnak az egyedei a lombhullás idejére elérik-e telelő pszeudopupa állapotot.
További vizsgálatok szükségesek az általunk feltételezett több nemzedék létezésének bizonyításához.
2024 augusztus második felétől kezdődően növekvő számban figyeltünk meg parazitoid darazsak által elpusztított liszteskék röplyukas pszeudopupáit A megfigyelő dobozban kikelt azonosítatlan Eretmocerus-faj imágóját lefényképeztük (2023.09.14.). 2020. október 10-én először észleltünk néhány, azonosítatlan mikrogombával fertőzött pszeudopupát, majd hasonló tüneteket 2024 októberében is megfigyeltünk.
A felhasznált fényképek a kártevők gyűjteményes fényképezése során (5K rendszer) készültek, az egyéb források a képek alatt kerültek megnevezésre.
Köszönetnyilvánítás:
Köszönettel tartozom Sütöriné dr. Diószegi Magdolna docens asszonynak (MATE TTDI Dísznövénytermesztési és Dendrológiai Tanszék), a Lonicera standishhi – télizöld lonc, a Lonicera maackii – bordáskérgű lonc (Budai Arborétum) fajok meghatározásáért.
Felhasznált irodalom:
- Balás, G. – Sáringer, G. (1983): Kertészeti kártevők Akadémiai Kiadó, Budapest 1982
- Bartha D. (2020): Fekete Lista. Magyarország inváziós fa- és cserjefajai. / Black List. Invasive tree and shrub species of Hungary. * Szürke Lista. Magyarország potenciálisan inváziós fa- és cserjefajai. / Grey List. Potentially invasive tree and shrub species of Hungary. – Soproni Egyetem Kiadó / University of Sopron Press, Sopron, 84 pp. Az interneten elérhető.
- Bethke,J. A.; Paine, T. D.; Nuessly, G. S. (1991): Comparative Biology, Morphometrics, and Development of Two Populations of Bemisia tabaci (Homoptera: Aleyrodidae) on Cotton and Poinsettia, Annals of the Entomological Society of America, Volume 84, Issue 4, 1 July 1991, Pages 407–411, https://doi.org/10.1093/aesa/84.4.407
- Bognár S. – Huzián L. (1979): Növényvédelmi állattan, Mezőgazdasági Kiadó, Budapest 1979.
- Evans, G.A. (2007) The Whiteflies (Hemiptera: Aleyrodidae) of the World and their Host Plants and Natural Enemies. USDA/Animal Plant Health Inspection Service. http://keys.lucidcentral.org/keys/v3/whitefly/PDF_PwP%20ETC/world-whitefly-catalog-Evans.pdf
- Gabrilov-Zimin, I.A., & Borisov, B. A. (2020): Aleuroclava aucubae (Homoptera: Aleyrodinea), a new adventive species for Russian Black Sea Coast, and its concomitant entomoparasitic fungus Conoideocrella luteorostrata (Ascomycota: Hypocreales: Clavicipitaceae), Zoosystematica Rossica 29 (1):3-10
- Hári, K., Fail, J, Streito, JC, Fetykó, K., Szita É., Haltrich. (2021): First record of Aleuroclava aucubae (Hemiptera Aleyrodidae) in Hungary, with a checklist of whiteflies occurring in the country, Redia
- Jermy, T. – Balázs, K. (szerk.): A növényvédelmi állattan kézikönyve I-VI. kötet Akadémiai Kiadó, Budapest, 1988-96
- Kanmiya, K. (2005): Mating Behaviour and Vibratory Signals in Whiteflies (Hemiptera: Aleyrodidae). in Insect sound and communication: Physiology, Behaviour, Ecology, and Evolution (CRC Press, 2005).
- Kontschán, J. (2023): Egy kevéssé ismert idegenhonos liszteske, (https://magyarmezogazdasag.hu/2023/05/19/egy-kevesse-ismert-idegenhonos-liszteske/)
- Lőrinczi, G., Torma, A. (2020): Állatrendszertan I.: Bazális csoportok (Non-bilateria) és ősszájúak (Prostostomia), Egyetemi jegyzet, SZEGED, 2020, Eletronikus kiadás
- Martin, J. H., Mifsud, D. & Rapisarda, C., (2000): The whiteflies (Hemiptera: Aleyrodidae) of Europe and the Mediterranean Basin. Bulletin of Entomological Research, 90: 407-448.
- MARTIN J.H., MOUND L.A., (2007): An annotated check list of the world’s whiteflies (Insecta: Hemiptera: Aleyrodidae). – Zootaxa, 1492: 1-84. https://doi.org/10.11646/zootaxa.1492.1.1
- Móczár, L. (1969): Állathatározó I. 338—340.p. Tankönyvkiadó, Budapest
- MOUND L.A., HALSEY S.H., (1978): Whitefly of the World. A systematic catalog of the Aleyrodidae (Homoptera) with host plant and natural enemy data. British Museum (Natural History)/John Wiley & Chichester, 340 pp. https://doi.org/ 10.5962/bhl.ti-tle.118687
- Patel C., Srivastava R.M., Samraj J.M. (2022): Comparative Study of Morphology and Developmental Biology of Two Agriculturally Important Whitefly Species Bemisia tabaci (Asia II 5) and Trialeurodes vaporariorum from North-Western Himalayan Region of India. Brazilian Archives of Biology and Technology. 2022, Vol.65, (https://www.scielo.br/j/babt/a/S4Xkskc7Q86DHyQxFyQGsrv/#)
- Penny J. Gullan, Jon H. Martin, in Encyclopedia of Insects (Second Edition), 2009
- Seljak, G. (2012): Six new alien phytophagous insect species recorded in Slovenia in 2011. – Acta Entomologica Slovenica, 20 (1): 31-44.
- Tóth, J. (szerk.) (2014): Erdészeti rovartan, Agroinform Kiadó, Budapest, 2014.
- VISNYA A., 1941 – Vorarbeiten zur Kenntnis der Aleurodiden-Fauna von Ungarn, nebst systematischen Bemerkungen über dei Gattungen Aleurochiton, Pealius und Bemisia (Homoptera). – Fragmenta Faunistica Hungarica, Suppl. 4: 1-20.





